第一百四十三章 亲缘与利他的关系
在此,有必要先行澄清某些精巧而荒谬的“科学社会学”理论。
【从“劳动创造了人”,到“剩余劳动”造成整个人类社会的异化结构,这个不真实的故事居然构成马克思主义社会学宏伟无比的基础框架。所谓“手不仅是劳动的器官,而且是劳动的产物”(引自《劳动在从猿到人转变过程中的作用》 恩格斯著),不过是在重复拉马克的“获得性遗传学说”的错误,其区别仅仅在于用“人类的手”置换了“长颈鹿的脖子”而已。实际的情形是:
a.任何后天获得的性状均不能通过生物遗传予以表达。也就是说,劳动──或曰“生物以其自体性状达成自存的行为之总和”──不创造任何生物遗传素质,反倒是生物遗传(以及在分子水平上所发生的生物变异)的自然素质决定着“生物劳动”的具体方式,就像鞭毛虫的辛劳并不是鞭毛虫得以产生和进化的动因一样。
b.任何生物均具有“剩余劳动”的潜能,可谓之“生物功能储备”。而且,愈原始的物种,其“剩余量”反而愈大,因此,原核单细胞绝不会发生体表营养膜上的紧张,反倒是真核单细胞越来越显得难以独存;不知蚂蚁能有多少可资榨取的“剩余劳动”,但它们却早已跨入了血腥、不义而分工有序的“奴隶社会”(可参阅达尔文在《物种起源》一书的〈特种本能〉章节中对“蚂蚁蓄奴”现象的生动描述);狮子每天尽可以懒睡二十个小时,但它们却不是以此为基础才建立起狮子社会的“等级压迫关系”的;回首今天的人类,实在忙之不迭,几至废寝忘食,且呈现紧张加剧的趋势,然而他们之间的相互“剥削”同样刻不容缓,绝不会因为“剩余劳动递减”而宣告大家散伙。
总之,这个理论体系中经不起推敲的东西太多,令人难以一一匡正。鉴于它是一个世纪以前的无知,故不复评述。】(1)
用“亲缘选择理论”来解释动物群体的利他性社会组合,是社会生物学建立其生物社会理论的核心。概括言之,即:自然选择的基本单位是基因;“自私的基因”(道金斯命题)倾向于在基因库中通过增殖复制以扩展自身的存在,由此造成同种DNA片段可以同时分布于许多不同的个体之内;如果某型基因既能够自私地抢占遗传上的优势,又赋有某种维护同型基因的利他品格,则它大抵会处于更有利的扩张地位(“利他”是“自私”的更高表现形式);从遗传学上讲,近亲之间具有较多的共同基因,亲缘个体之间的利他主义关系,就是基因的这种共同利益之体现。
为此,人们经常借用汉密尔顿模式(亦称“基因亲缘关系的利他行为损益模式”或“基因利他主义模式”)来表述生物种群的社会关系:
C表示“基因支出或折损”,B表示“基因补偿或收益”,r表示生物个体之间的“亲缘关系系数”。从这个公式的推导中可以看出,当个体的直接增殖效果降低一个C量级时,如果它同时能够从旁系亲属中使其基因遗传增加一个相等的r·B量级,即
C=r·B
则要么r值必须很大,要么B值就得远远超过C值,否则利他性行为势必呈现出一个亏损结果,自然选择就有可能淘汰这些优雅高尚的利他主义基因。然而十分不幸的是,除了同卵双胞胎或无性繁殖之外,r值总是<1,因为在二倍体生物──绝大多数动物和植物包括人类在内都属此型──的繁殖系列上,父本和母本各以减数分裂的形式在其精卵生殖细胞中贡献出恰好半数的单倍体组基因,子代必须以基因重组的方式重新形成一个二倍体基因组个体。因此,子代与亲代共有1/2的同型基因,且按倍数关系隔代递减,即直系祖孙之间共有1/4的相同基因,直系曾祖孙之间共有 1/8的相同基因,依此类推。旁系亲属之间的横向隔层关系亦然。难怪群体遗传学家J·B·S霍尔丹曾开玩笑说,他会甘愿冒险跳进水里去救三个溺水的同胞兄弟或九个溺水的堂表兄弟。因为出于对自然选择的尊重,他必须事先盘算停当,他与亲兄弟共有1/2的基因数,而与堂表兄弟共有1/8 的基因数,只有超过这个分母量级以上时,他才不至于让自身可贵的基因亏本。倘若溺水的亲兄弟只有一个,或溺水的堂表兄弟只有七个,再或者,纵然遇险的非亲非故者以数十人众计,他也应该只好作壁上观了。
不过,从表面上看,恰恰是这个理论最圆满地阐明了生物亲缘之间的利他行为关系。仍以蜜蜂社会为例:蜂群基本上是一个雌体社会,如果单从基因组型的角度考察,它们之间的互助程度与其基因同型率存在着某种相关关系。跟其他物种不同,雄蜂没有父亲,它们是由未受精卵发育而成的单倍体基因载体。在母蜂生殖系统的解剖结构上,中输卵管的上方有一个直径约1.5毫米的贮精球,贮精球内贮藏交配所得的雄蜂精液,供母蜂一生使用,贮精球管口由肌肉收缩来控制精液的排放,使卵子受精或不受精。受精卵发育成同为雌体的二倍体工蜂或母蜂(蜂王)。母蜂与工蜂的表型差别由后天发育条件的相异所致,与基因组型无关。因此,蜂群的基因谱系别具一格:作为子女的雄蜂和雌蜂与其母本仍然保持1/2 的共同基因,但一个雌体与其姐妹所共有的基因呈现3/4的高比率,即来自父亲的全部单倍体基因以及来自母亲二倍体基因中的1/2;而与其兄弟所共有的基因呈现1/4的低比率,因为她和他们只有共同的母亲,所以结果与同母异父的子女之间的情形相似;反过来看,雌性子代之间的共同基因高于与其亲代的亲缘关系系数;或者,蜂王登基伊始,同代的作为子民的工蜂们在基因上与她一律处在平等的关系中;而雄性子代与其势力强大的姐妹们之间的共同基因却跌落到与其母亲的关系之下了。果然,实际的社群共处关系也相应分化开来:工蜂们为了以雌性为主的群体利益不惜欣然赴死,因为它们做出的牺牲可以凭借高比值r获得较高的B值补报,与其把它们的献身精神看作是对蜂王的效忠,可能不如视为是对姐妹们的维护更确靠;而雄蜂受到的待遇就大为不同了,一旦过了交配繁殖的季节,尤其当外界蜜源稀少的时候,工蜂们就把既不会采食也无能自卫的雄蜂兄弟赶出巢门,听任其自生自灭而绝不以恻隐之心为怀。社会生物学家特里弗斯和黑尔的研究还发现,在同属此类基因谱系的某种膜翅目黄蚁社群中,“雌性的生物量(按重量估计)比雄性的生物量高3倍,因此(可以认为),工蚁喂养前者要比喂养后者多付出3倍的精力。”(《科学》杂志,1976年第191期第249页,转引自《社会生物学》,米歇尔·弗伊 著,殷世才 孙兆通 译。)这个倍数关系与它们之间的基因同型比率恰好吻合。【而且,要探讨生物亲缘之间的天然亲情倾向及其渊源,也没有比这个理论更好的解释了。有兴趣的读者可参阅E·O ·威尔逊的著作《社会生物学──新的综合》以及其他学者的有关专著。】
然而,如果再从体质性状分化整合的批判角度予以考察,则会发现用基因亲缘损益模式来界定膜翅目族类的社会关系实在是破绽百出:工蜂们生就一副从事经济和御敌的职业性躯体和器官,为了自存,也为了自存必需的社会繁荣,她们反而更殷勤地侍奉基因同型率(即亲缘关系系数)明显低于工蜂姐妹的上辈蜂王(蜂王的存活期通常是工蜂寿限的数倍以上);而且,万一蜂王缺失,举“国”不宁,你于此时可做一个实验,从另外的蜂群中诱入一只母蜂,虽说血缘迥异,工蜂们的殷勤之状不改,照例尊以王礼,且即刻可以恢复蜂王国的旧有秩序;再者,由此外来蜂王产卵羽化的工蜂可能与本群中原来的工蜂们全无姐妹之间的基因关系,但它们之间的亲密互助情谊一如同母所出,断不会发生种群社会内的基因宗派勾斗;在社会形势稳定的“分蜂间期”,体质未衰的母蜂会断然咬杀发育成王储的骨血千金,却同时可能恬不知耻地去与毫无亲缘关系可言的外族雄蜂调情戏淫;工蜂们历来严守巢门,不许任何外群的成员随便窜入,即便乞食的来客恰恰是从本群中分蜂而别的血亲姐妹也翻脸不予相认,但对可能成为“继父”(或“续弦姐夫”)的浪荡雄蜂却一反常态地开放门禁, 恭迎入室,且给其把盏敬饭, 一点也不因他食相凶狠而显出一丝一毫的嫌厌情绪;母蜂不免有衰老停育的一天,一般还等不到这一刻的来临,为了防止社会内乱,或者为了同一目的而需要促成分蜂(自然分群)的实现,工蜂们竟会毫不怜念老母的基因半值之生恩,硬是全然停尽赡养之孝道,逼得老母只好自去巢房取食,不止如此,原先的侍卫蜂此时还爬到女王身上抖动骚扰,迫使母蜂在蜂王台内产卵,然后精心护育幼主成长,羽化登基的新女王倘若不止一只,则“天无二日”的政治法则将驱使身为姐妹的年轻母蜂发生一场王位之争,且必欲置对方于死地而后快,其血腥之状俨如唐王朝开国初期的“玄武门之变”,此时颤巍巍的老母只好知趣地率其部分旧臣弃“国”亡命了。上述所有这些背离亲缘利他原则的“缺德”行径,有效地维护了异质整合社会的生机和秩序。在基因利他主义与社会组织分化之间发生矛盾之际,正是这些最能证明亲缘关系学说及其换算公式的同一物种,毅然抛弃了它们的天伦之情,反而采用最能证伪亲缘选择理论的行为方式,在“社会”的祭坛上,不惜决然供奉出血肉相连的基因牺牲品。而且,汉密尔顿模式“解释不了所有膜翅目社会,因为另一些社会昆虫群体──白蚁──具有严格的二倍体遗传”(引文出处同下),就是说,它们的亲缘关系系数r不可能形成3/4的高值;“此外还有一个问题,即膜翅目群有许多能生育的雌性(称为‘多雌群体’),或只有一只雌性,但接受多只雄性授精,这种例子很多。在这两种情况下,同一群里两只工蚁的平均亲属关系系数就会降到1/2以下。”(引自《社会生物学》,〔法〕米歇尔·弗伊 著,殷世才 孙兆通 译。)【变换一个视角来看,社会生物学所主张的基因同型率与利他性组合之间的相关关系,似乎只能用来阐释机体细胞的“体内社会化”现象:同一生物有机体的所有细胞确系同一基因组型,相应地,其结构性蛋白质的组成序列也完全一致,各细胞之间不仅利他互助,相依为命,而且抗原性无异(亦即完美无误地具有威尔逊所谓的识别性“绿须效应”),在生理情况下绝不会发生上述见于蜜蜂社会中的那些“失常性自残”之举。如果我们做一个组织相容性实验,即对任一多细胞高分化型的动物进行选定器官或组织细胞的移植手术,则受体动物的排异反应烈度,将严格依据它与供体动物之间的基因同型系数而呈现出从零排斥到完全排斥的直线相关关系,换一个浅显的说法,就是自体移植则相安无事,异体移植则亲缘关系愈远者可能表现出愈严重的排异反应后果。故对需要接受器官移植的实验动物或患者事先进行供体筛选,一般总要从直系亲属中就近考虑。很明显,从生物机体内的“社会关系”上看,汉密尔顿模式显得分外贴切,在这里,r系数获得了满足值的正整数1,任一C细胞的牺牲奉献必然等量换来另一B细胞的受益所得,而且随着r系数的任意变化,等号两边的损益增减始终保持相等。然而,这样一组规定毕竟仅仅表达了纯粹生物学上的某种生理机制,如前所述,它与体外真正的社会存在系统并不符合,社会生物学显然将这一公式用错了地方。】
我无意借此批驳或否认基因利他现象的客观存在,但我实在看不出基因的“自私”或者“利他”与社会构成有什么直接的、要紧的基础性关系。须知以“自私”或“利他”作为社会构成的“道德前提”早已是老生常谈了,但它什么也未能说明,因为“自私”和“利他”本身尚有待说明。实际上,所谓“自私”乃是任何存在物的原始“自在性”的变态表达;所谓“利他”乃是任何分化物的后衍“自为性”的具体贯彻;它只能用来显示物质属性演化或依存属性特化的一般轨迹,却不能视其为生物社会构成的特殊要素和基本动因。【如果基因分子的求存本性可谓之为“自私”,则任何物质的存在都是“自私”的,因为任何作为“在者”的物体都有一个不可让渡的自在前提,有如同一轨道上并行着两个以上运行速度不等的天体,它们要么撞合为一,要么击碎相遇者使之瓦解或逸出该轨道,其中任一天体都不会为了给其他天体让路而“利他”式地自我消灭。不过,天文学家绝不会用这种方式来图解天体运行,就像物理学家绝不会用电子与质子之间的“利他”关系来诠释原子构成一样。】(2)
实际的情形是,在膜翅目社会中,基因的亲缘关系系数与异质整合的结构化社会形态之间存在着某种交叉发展的偶合关系或一过性的重叠关系。也就是说,在泥盆纪以后的生物进化途中,造成社会性生物性状分化的代偿条件和机制尚处在可能借助于染色体倍数异化的阶段。这种说法可以得到如下事实的充分支持:即单倍体或奇数多倍体生物,既不见于原始单细胞生物,也不见于高度进化的正常脊椎动物或人类,它只出现于某些低等植物的配子体和个别昆虫的雄体中(单倍体组),以及某些无性繁殖的植物种类里(三倍体组)。然而,生物系统的社会演化,却以一种与基因拷贝的同型率全然无关的形式展开:起初,在原始单细胞生物阶段,由于增殖形态属于无性繁殖的简单分裂或出芽生殖,因而在其任一亲缘群体之中,所有个员的共同基因比率均为100%,即r系数为1,但在它们的初级社会里,你无论如何也找不到利他主义的行为迹象或性状配合,换言之,生命与社会初诞之际,所谓生物利他性社会关系的决定因素──基因亲缘系数或基因同型率,就与社会组合张力之间表现为根本脱节的失构状态;继之,在亲缘r系数明显下浮的二倍体两性繁殖的广大中级社会动物种群之间,其各自表征的社会整合度悬殊,即在基因亲缘系数大致相当的动物社会坐标系内,其利他性依存关系呈现出完全离散的无相关性分布状态,尽管如此,中级动物的利他性社会性状分化反倒在总体上显示出大幅攀升之动势却是不容置疑的事实;最后,再看整合度更高的人类晚级社会,其非亲缘化的历史发展趋势必然使群体成员之间的平均同型基因比率逐渐降至极度稀释的境地,然而恰恰是在这种情况下,生物社会的利他性行为和利他性结构正在阔步走向至高无上的境界。【从人类原始亲缘社会的“氏族”,到亲缘系数扩散性降低的“胞族”,再到多个胞族联合而成的“部落”乃至“部落联盟”,尔后进至中古代“民族”,再进至“多民族国家”以及当今已具雏形的“多国联盟”,这个社会结构扩大化和密合化的进程反而恰恰与“大家族”乃至“小家庭”的解体进程相伴而行,亦即恰恰与社群平均基因亲缘系数趋于下降的进程相互吻合。】
至此,除了膜翅目昆虫社会这个交叉点例外,基因亲缘系数与利他性社会组合之间基本上呈现为一目了然的偏离态势。其反向演运过程可用如下模式化的坐标图示之:
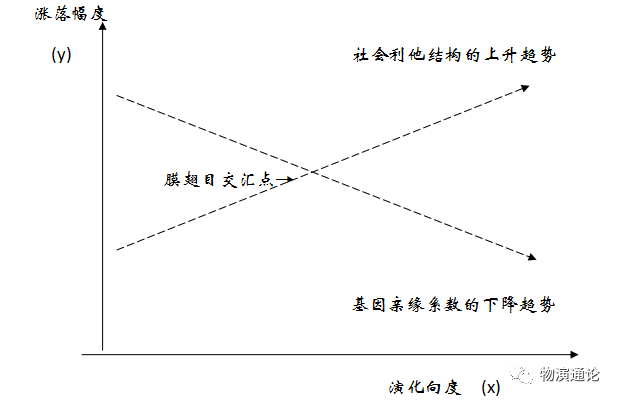
【注解:亲缘关系自将在一切生物社会乃至人类社会中永远存在下去,但它并不是生物利他性社会构成的基本动因。自初级社会到中级社会的某一位点──如膜翅目昆虫社会,基因r系数趋降,社会利他结构趋升,二者的逆向演动恰好交汇在膜翅目社会的分化形态上,使之表现出一定程度的粗略相关;过后,这两条运动轨迹继续沿着各自的原定方向在生物社会衍存系统中运行,结果却造成利他性社会分化日益背离基因亲缘系数的局面。】(3)
综上所述,我们可以对本章之讨论给以这样一个扼要的总结:“基因决定论”乃是威尔逊(E·O·Wilson )创立社会生物学的基本命题和出发点,他那句振聋发聩的著名论断:“有机体只是DNA制造更多DNA的工具”,着实一举将生物存在以及社会存在全然放置在与分子存在同一的自然脉络上了。不过,这里遇到一些逻辑上的麻烦:如果DNA是生物层位的基底规定者(这一点已为现代分子生物学所证实),则它就不可能同时又是社会层位的直接规定要素,否则,生物与社会自属同一代偿衍存位格上的同一物类,亦即二者处于无可区分的逻辑同格上,这不啻从根本上否定了社会存在。再者,如果继续往前追溯的话,可以说“DNA无非又是有机小分子、原子乃至基本粒子的衍存工具”,然则是否可以得出如此结论:一切后衍物相均可使用原子物理学的方法来研究呢?【显然,全部的问题发生在这样一个关节上:当分子存在(如DNA)以某种特定方式支配着生物存在之时,生物存在又将以何种特定方式支配社会存在?所谓“特定方式”, 其实就是“自然衍存位相各运动质点上的特定代偿属性”之别称。即是说,针对任一具体物相而言,你必须找到切近于它的具体决定因素或曰“直接代偿规定”(因为世上万物之间的“间接依存关系”是无穷无尽的);而要做到这一点,你又必须首先找到整个自然衍存流程的统一规定;一望而知,用“基因观点”来讨论“社会物相”是对上述二者一概失察的表现。】(4)
注释:
(1)本章的主要内容是澄清既往“科学社会学”的错误,如马克思主义社会学的理论基础就具有典型的错误:a、“劳动创造了人”、“手不仅是劳动的器官,而且是劳动的产物”是已经被证明错误的拉马克“获得性遗传学说”的翻版。因为生物后天获得的性状并不改变基因序列,如运动员练出了最发达的肌肉不可能通过基因遗传给孩子,数学家的后天认知也不可能通过基因遗传给孩子,孩子要想实现和父辈一样的成就,也需要进行后天的训练和学习。显然,劳动也不会改变你的基因序列。b、“剩余劳动”和社会剥削没有关系。所谓“剩余劳动”,追究到底,就是“生物功能储备”,而随着生物的进化,其“生物功能储备”是递减的。单细胞生物的“生物功能储备”几乎是无限的,但阶级划分与剥削无由发生。人类社会中个体的“生物功能储备”其实是最有限的,更不是社会产生阶级和剥削的原因。阶级的分化归根到底是自然分化在人类社会中的延续,后续第一百七十章有详细的论述,此处不赘述。
(2)澄清既往社会生物学上“基因决定论”的错误。本注释只是有重点的说明和概述,书中大量生物现象的例证与详细说明注释中不再重复,可通过看书去完整地了解。
“基因决定论”下生物种群社会关系的汉密尔顿模式(亦称“基因亲缘关系的利他行为损益模式”或“基因利他主义模式”)表达为:B/C=1/r( C表示“基因支出或折损”,B表示“基因补偿或收益”,r表示生物个体之间的“亲缘关系系数”)。它可以解释膜翅目社会的部分利他行为,但有更多的利他行为无法靠这个公式解释。所以,汉密尔顿模式仅仅表达了纯粹生物学上的某种生理机制,其基因同型率与利他性组合之间的相关关系在器官移植手术中充分表达,但它并不符合更高层级的社会代偿中利他行为的解读。如在单细胞生物社会,亲缘关系系数r=1,C=r·B,哪怕以我一个细胞的死换其他任一细胞的活都是等值的,不亏本,而利他程度越高,即能换活的细胞越多,基因收益越大。但很显然,单细胞生物社会并无半点积极的利他行为。再如中级社会,社会结构度越高,亲缘关系越离散,社会的利他程度越高,越是需要通过服务别人来获取自身的生存。
所以,基因利他行为的客观存在并不能构成解读自私和利他的原因。自私和利他是由更深在的存在度决定的:存在度高的存在自在而圆满,无需利他或害他,如单细胞生物相对后衍生物存在度最高,独自即可存活,自然无需和其他细胞发生利害关系;存在度低的存在必须通过自为的利他才能换取自身的依存条件,如人类晚级社会的社会分工下,人人都需要通过服务于他人、服务于社会才能换取到自身需要的生存条件。
(3)汉密尔顿模式之所以能解释膜翅目社会的部分利他行为,是因为生物社会是社会化进程中,基因亲缘系数趋降,社会利他结构趋升,膜翅目社会刚好是这两种趋势的交叉点。从寒武纪前后的“显生现象”到进入泥盆纪以后某个特定阶段,中级生物社会内部体质性状差异导致的代偿方式或耦合方式是通过性分化下的染色体倍数分化来实现的,明显的例子就是单倍体组只见于低等植物的配子体和个别昆虫(如蜜蜂)的雄体中,奇数多倍体组只见于无性繁殖的植物中,而主流的中级社会都是双倍体。蜜蜂社会正处于这个代偿位相上,所以染色体倍数异化(即雄蜂是单倍体、雌蜂和蜂王是双倍体)导致了蜜蜂间的性状差异及其不同的代偿方式,也就是说,蜜蜂利他的耦合方式(即代偿方式)正好与染色体倍数分化相关,由此达成了基因亲缘系数与利他在一定程度上能匹配。但主流的社会分化显然与基因亲缘系数或基因复制的同型率无关:如单细胞生物社会基因复制的同型率最高(r=1),反而最没有利他行为;中级社会中基因复制同型率大致相当的动物社会,其利他行为并不一致,反而呈现为离散的不相关状态,也就是说,基因复制的同型率和利他行为无关。决定利他行为的是主体的存在度或分化程度,所以,人类晚级社会以后,人类社会逐渐从亲缘社会变成非亲缘社会,社会中的利他行为却越来越成为社会分工下的必须。也就是说,物演的生物社会化进程中,基因亲缘系数趋降,从单细胞生物社会的基因亲缘系数为1到异性繁殖下的50%,到人类从亲缘社会扩展为非亲缘社会;社会的利他性代偿趋升,从单细胞生物的利他为零,到中级社会社会成员间的较低程度的功能分化,到人类文明社会化的大分工;所以,基因亲缘系数和社会的利他性代偿一直是彼此偏离的。
换句话说,既往的社会学对于动物包括人的利他行为展开了很多讨论,但没有一个令人信服的明确的结果,所以需要重新展开讨论:什么是利他?利他的本质是什么?
首先,在递弱代偿的衍存律下一切代偿都是为求存服务的,无论高存在度的自在表现还是低存在度的自为表现自然也不例外。自在其实就是靠自己满足求存的自私。自为其实就是在失位的社会中找到自己的社会依存定位,通过与他人合作共同实现整个社会的求存进而满足自己求存,即为了自存的社会利他行为。哪怕是竞争也是基于特定范围资源不足所寻求的特定资源重新分配的代偿方式。如前衍的无机存在是无需竞争的,最低等的生命单细胞生物也是因为超量繁殖到资源不足才会引发竞争,哪怕是中级社会时,根据血缘群团划分领地,因为领地够大、资源够丰富,同类种群之间并不发生竞争关系,只有当资源紧张,比如随着智人迁徙人类占满地球再无法通过迁徙解决资源问题时,血缘群团或氏族社会之间的竞争就不可避免。因为每个氏族社会的力量差距不是很大,为了胜利就会需求与其他氏族的联合,联合的规模越大力量就越大,就越有绝对的胜利把握,于是便有了从氏族联合成部落、部落联盟、城邦或王国、封建君主国家、议会民主国家、国家联盟、跨洲联盟乃至人类命运共同体的社会结构化进程。这个进程就是合作利他共同求存的社会分化耦合进程。无论社会成员还是社会整体,残化程度越来越大,利他程度(即耦合需要)就越来越大。所以,利他的本质就是不同衍存位相上存在者的耦合代偿方式。但耦合后的结构存在更复杂更脆弱、耗费资源更多,进一步导致资源不足,于是恶性循坏至对人类命运共同体的追求,这需要更高层级、更大范围的利他行为。如中国对非洲的援助,也有自己的经济和政治目的,希望达成与非洲更多的互助与合作,建立更大的分化耦合结构。
其次,具体来看,社会层级的利他本质直接就是社会特定质态下的耦合代偿方式,确切而言,就是生存性状分化耦合的代偿方式,通俗而言,就是求存行为分化耦合的代偿方式或社会分工合作的代偿方式。
所以,一旦弄清了利他的本质,就自然否定了基因决定论,也澄清了自私与利他的关系。如果说自私是满足自己求存,那么利他是通过满足社会整体求存而实现自我求存。用书中的话说:所谓“自私”乃是任何存在物的原始“自在性”的变态表达;所谓“利他”乃是任何分化物的后衍“自为性”的具体贯彻;它只能用来显示物质属性演化或依存属性特化的一般轨迹,却不能视其为生物社会构成的特殊要素和基本动因。所谓“利他性”其实并不与基因的同型系数相干,你的残化使你无由自私或自在;你要是圆满,你想利他还利他不成。自私是自斥于群化结构之外的一种完善;利他是自私不得的一腔无奈。你残化了,你以利他换取自私的前提,你的利他主义其实完全是自私的变种。从常识的角度来看,利他主义行为不过是客观上更为狡猾的自私的同一;从哲学的角度来看,利他主义行为无疑是自然存在物从“低依存度的自在”流向“高依存度的自为”之必然。换言之,自私意味着自私者(或“自在者”)融不进社会的结构,应该说,这不是由于它的自私,而是它根本就不需要这种结构。如果你需要这种结构以为补缺,你还硬以自私与之相抗,处处显得格格不入,那你真是自取灭亡。总之,无论是情态上或意态上的自私和利他,都不能成为社会整合结构的基础和原因,反过来说倒不会出什么大错:恰恰是社会成员与社会构态的演化进程,规定着“自私”或“利他”的生物行为方式。
毋容置疑,从整个社会的结构化进程来看,利他只是个体在失位社会状态下的定位依存方式。其求存本质和单细胞生物社会中每个细胞无需利他而自在求存(或自私求存)并无区别。这是站在最深刻的个人都是社会人的角度或关系来说明的。表面上,郑也夫把利他行为分成三种:第一、本能的利他,如母爱(这和基因的自然选择有关);第二、有利益相关的利他如通过职业利他获取个人求存,如通过个人捐款打造个人声誉;三、无关利益的利他,如给乞丐钱、给流浪猫食物、做好事不留名等。这些利他,归根到底,都是维护社会整体求存的:亲缘社会以性耦合为中心,就是体质分化下的耦合求存或定位依存方式,这种社会环境下的母爱和血缘之爱是社会得以维系的基础。非血缘社会的人类晚级社会,所有和利益相关的利他行为都是以维护他人利益、实现社会整体求存的方式地维护自己的求存,包括维护自己在社会中的身份和地位也是为了更好的求存。和利益无关的利他,甚至舍己为人的利他要么是家庭、社会后天文化教育使然(文化也是维护社会和谐或整体求存的),能自觉同类乃至万物的一体求存关系;要么是亲缘社会中的恻隐之心在非亲缘社会中继续延续(越基础的代偿越具有奠基性、决定性、稳定性和普遍性),或者在意识深层不自觉地有着同类乃至万物的一体求存感性的模糊的认知。比如当下有人提出生态主义的概念,指出人类下一期文明的任务就是维护好地球生态的良性循坏,这仿佛超越了人与人之间的利他,是一种更高境界的利他,但它的本质还是维护人类的求存,因为人也是万物之一,自然生态是其最底层的依存条件,万物的衍存关系决定了万物一体的相关性。所以,无论人类的哪种利他行为,其代偿而出就一定有着维护社会整体求存进而维系个体求存的效用。
进一步而言,如果把“利他”视为“善”,把“自私”视为“恶”,则善恶一元,都基于依存的需要。万物都是本原存在的分化物,万物一体或万物的统一性的最深直觉都是“先验的沉淀在意识深层中的无意识基底”,这是人性中的神性来源,也是古人说“至善无善”(即善而不以为善)的道理;但切近的依存现实中,繁殖量远大于生存量的资源限制导致了竞争,这是人性中的兽性来源。极端的矛盾导致极端的兽性。正常情况下人都是平衡的,极端情况下才需要焕发出神性或兽性。
关于“善”,第一百七十四章讲道德的时候也会进一步阐述,这里不再赘述。
由此可见,社会性利他行为和个体求存的自私并无矛盾,而且,社会性利他行为正是个体在社会中实现个体依存的求存方式——也就是自私的方式。
(4)总之,生物体质层面或生理层面的代偿(即体内的细胞社会化)和生物的社会性代偿(即体外的生物社会化)是不同的代偿层级,其具体的代偿方式肯定有所区别。这就是如同物演进化中不同的代偿层级具体的代偿方式必有所区别:原子进化是物理现象,遵循具体的物理规律;分子进化是化学现象,遵循具体的化学规律;生物进化是生物现象,遵循具体的生物规律。当然,不同层级的代偿规律只是其特定层级的表观现象,在整个物演序列的进程中,递弱代偿的存在性才是共同的终极本质。所以,当我们寻求对某种现象的解释时,我们首先要把它放在最大的整体上,找到最大整体的统一规定,即自然衍存的统一规定,其次再针对该存在者的具体衍存位相去寻找该位相下或该代偿层级下的具体决定要素。很显然,用基因决定论来解读社会的利他行为包括具体的膜翅目社会的利他行为必然是错误的,就因为它既不理解自然衍存的总规律,也没找到中级社会以及具体的膜翅目社会利他行为的直接代偿规定。
换句话说,对膜翅目利他行为和亲缘关系的探讨首先需要把利他行为放在整个社会的结构化进程中去看,才能看清利他行为随着社会化进程趋升的总趋势,与之相悖的基因亲缘系数趋降的总趋势自然不能成为利他行为的决定要素。于是,膜翅目社会中能用基因亲缘系数解读的利他行为就只能是随着社会结构化进程利他行为趋升和基因亲缘系数趋降的一过性重叠关系。这时候要说明具体的膜翅目利他行为和亲缘关系就转化为膜翅目社会化分工与亲缘的关系,这就是膜翅目特有的靠种群内部的体质分化来完成膜翅目的社会化的分工与合作,具体而言,就是染色体倍数的分化决定了膜翅目社会种群内部的体质多样性,从而决定了膜翅目的社会化的分工与合作,于是,这种和基因密切相关的社会分工就导致了基因与利他关系的误会,出现基因决定论。
注:本章以澄清既往社会学的错误为目的,主要澄清了膜翅目社会中基因亲缘系数与社会利他行为的关系,从而否定了社会层面的基因决定论,以导出下一章对社会的直接决定要素的探寻。
(文中配图来自网络)
…………………………
希望本公众号对您学习《物演通论》有帮助!思想的传播需要我的初心和坚持,也需要您的支持和鼓励!欢迎提供反馈,或赞赏、转发,或批评、指正。谢谢!
1、欢迎读书困难的同学关注“物演通论注释”公众号,有逐章的注释以供参考,二维码如下:

2、请关注“物演通论看世界”公众号,二维码如下:

3、请关注“物演通论读书会”公众号,二维码如下:

